Parasaurolophus
From Wikipedia, the free encyclopedia
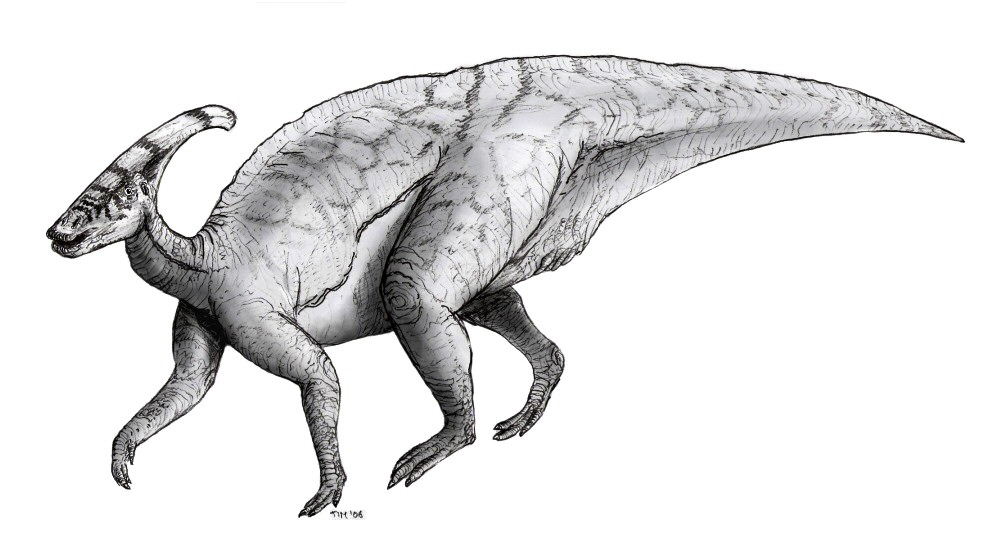
[Photo] Sketch of the hadrosaurid Parasaurolophus in tetrapod or quadrupedal pose, anatomically correct. Made by Tim Bekaert, 2006. Like other hadrosaurs, Parasaurolophus was facultatively bipedal, i.e. they could alternate between two legs and four, probably preferring a quadrupedal gait while they foraged for food and assuming a bipedal mode for faster running.
Parasaurolophus (meaning "near crested lizard" in reference to Saurolophus; pronounced RP [??pæ????s??ː??????l??f??s], GA [??pæ????s????????l??f??s], in common usage is also RP [??pæ??????s??ː??????l????f??s], GA [??pæ??????s????????lo??f??s]) is a genus of ornithopod dinosaur from the Late Cretaceous Period of what is now North America, about 76-73 million years ago. It was a herbivore that walked both as a biped and a quadruped. Three species are recognized: P. walkeri (the type species), P. tubicen, and the short-crested P. cyrtocristatus. Remains are known from Alberta (Canada), and New Mexico and Utah (USA). It was first described in 1922 by William Parks from a skull and partial skeleton in Alberta.
Parasaurolophus is a hadrosaurid (duckbilled dinosaur), part of a diverse family of Cretaceous dinosaurs known for their range of bizarre head adornments. This genus is known for its large, elaborate cranial crest, which at its largest forms a long curved tube projecting upwards and back from the skull. Charonosaurus from China, which may have been its closest relative, had a similar skull and potentially a similar crest. The crest has been much discussed by scientists; the consensus is that major functions included visual recognition of both species and gender, acoustic resonance, and thermoregulation. It is one of the rarer duckbills, known from only a handful of good specimens.
Description
As is the case with most dinosaurs, the skeleton of Parasaurolophus is incompletely known. The length of the type specimen of P. walkeri is estimated at 9.5 meters (31 ft). Its skull is about 1.6 meters (5.2 ft) long, including the crest, whereas the type skull of P. tubicen is over 2.0 meters (6.6 ft) long, indicating a larger animal. Its weight is estimated at 2.5 tonnes (2.7 tons). Its single known forelimb was relatively short for a hadrosaurid, with a short but wide shoulder blade. The thighbone measures 103 centimeters (3.38 ft) long in P. walkeri and is robust for its length when compared to other hadrosaurids. The upper arm and pelvic bones were also heavily built.
Like other hadrosaurids, it was able to walk on either two legs or four. It probably preferred to forage for food on four legs, but ran on two. The neural spines of the vertebrae were tall, as was common in lambeosaurines; tallest over the hips, they increased the height of the back. Skin impressions are known for P. walkeri, showing uniform tubercle-like scales but no larger structures.
The most noticeable feature was the cranial crest, which protruded from the rear of the head and was made up of the premaxilla and nasal bones. The P. walkeri type specimen has a notch in the neural spines near where the crest would hit the back, but this may be a pathology peculiar to this individual. William Parks, who named the genus, hypothesized that a ligament ran from the crest to the notch to support the head. Although this idea seems unlikely, Parasaurolophus is sometimes restored with a skin flap from the crest to the neck.
The crest was hollow, with distinct tubes leading from each nostril to the end of the crest before reversing direction and heading back down the crest and into the skull. The tubes were simplest in P. walkeri, and more complex in P. tubicen, where some tubes were blind and others met and separated. While P. walkeri and P. tubicen had long crests with only slight curvature, P. cyrtocristatus had a short crest with a more circular profile.
Classification
As its name implies, Parasaurolophus was initially thought to be closely related to Saurolophus because of its superficially similar crest. However, it was soon reassessed as a member of the lambeosaurine subfamily of hadrosaurids???Saurolophus is an hadrosaurine. It is usually interpreted as a separate offshoot of the lambeosaurines, distinct from the helmet-crested Corythosaurus, Hypacrosaurus, and Lambeosaurus. Its closest known relative appears to be Charonosaurus, a lambeosaurine with a similar skull (but no complete crest yet) from the Amur region of northeastern China. P. cyrtocristatus, with its short, rounder crest, may be the most basal of the three known Parasaurolophus species, or it may represent subadult or female specimens of P. tubicen.
Discovery and naming
Meaning "near crested lizard", Parasaurolophus' name is derived from the Greek para/παρα "beside" or "near", saurus/σαυρο?? "lizard" and lophos/λοφο?? "crest". It is based on a skull and partial skeleton missing most of the tail and the hind legs below the knees (ROM 768), which was found by a field party from the University of Toronto in 1920 near Sand Creek along the Red Deer River in Alberta, Canada. These rocks are now known as the Campanian-age Upper Cretaceous Dinosaur Park Formation. William Parks named the specimen P. walkeri in honor of Sir Byron Edmund Walker, Chairman of the Board of Trustees of the Royal Ontario Museum. Parasaurolophus remains are rare in Alberta, with only three other specimens from the Dinosaur Park Formation, all lacking skulls, possibly belonging to the genus.
In 1921, Charles H. Sternberg recovered a partial skull (PMU.R1250) from what is now known as the slightly younger Kirtland Formation in San Juan County, New Mexico. This specimen was sent to Uppsala, Sweden, where Carl Wiman described it as a second species, P. tubicen, in 1931. The specific epithet is derived from the Latin t??b??c??n "trumpeter". A second, nearly complete P. tubicen skull (NMMNH P-25100) was found in New Mexico in 1995. Using computed tomography of this skull, Robert Sullivan and Thomas Williamson gave the genus a monographic treatment in 1999, covering aspects of its anatomy and taxonomy, and the functions of its crest. Williamson later published an independent review of the remains, disagreeing with the taxonomic conclusions.
John Ostrom described another good specimen (FMNH P27393) from New Mexico as P. cyrtocristatus in 1961. It includes a partial skull with a short, rounded crest, and much of the postcranial skeleton except for the feet, neck, and parts of the tail. Its specific name is derived from the Latin curtus "shortened" and cristatus "crested". The specimen was found in either the top of the Fruitland Formation or, more likely, the base of the overlying Kirtland Formation. The range of this species was expanded in 1979, when David B. Weishampel and James A. Jensen described a partial skull with a similar crest (BYU 2467) from the Campanian-age Kaiparowits Formation of Garfield County, Utah. Since then, another skull has been found in Utah with the short/round P. cyrtocristatus crest morphology.
Species
The type species P. walkeri, from Alberta, is known from a single specimen. It differs from P. tubicen by having simpler tubes in its crest, and from P. cyrtocristatus by having a long, unrounded crest and a longer upper arm than forearm.
P. tubicen, from New Mexico, is known from the remains of at least three individuals. It is the largest species, with more complex air passages in its crest than P. walkeri, and a longer, straighter crest than P. cyrtocristatus.
P. cyrtocristatus, from New Mexico and Utah, is known from three possible specimens. It is the smallest species, with a short rounded crest. Its small size and the form of its crest have led several scientists to suggest that it represents juveniles or females of P. tubicen, which is roughly the same age and from the same formation in New Mexico. As noted by Thomas Williamson, the type material of P. cyrtocristatus is about 72% the size of P. tubicen, close to the size at which other lambeosaurines are interpreted to begin showing definitive sexual dimorphism in their crests (~70% of adult size). This position has been rejected in recent reviews of lambeosaurines.
Paleoecology
Parasaurolophus walkeri, from the Dinosaur Park Formation, was a member of a diverse and well-documented fauna of prehistoric animals, including well-known dinosaurs such as the horned Centrosaurus, Styracosaurus, and Chasmosaurus; fellow duckbills Prosaurolophus, Gryposaurus, Corythosaurus, and Lambeosaurus; tyrannosaurid Gorgosaurus; and armored Edmontonia and Euoplocephalus. It was a rare constituent of this fauna. The Dinosaur Park Formation is interpreted as a low-relief setting of rivers and floodplains that became more swampy and influenced by marine conditions over time as the Western Interior Seaway transgressed westward. The climate was warmer than present-day Alberta, without frost, but with wetter and drier seasons. Conifers were apparently the dominant canopy plants, with an understory of ferns, tree ferns, and angiosperms.
The New Mexican species shared their environment with the large sauropod Alamosaurus, duckbill Kritosaurus, horned Pentaceratops, armored Nodocephalosaurus, Saurornitholestes, and currently unnamed tyrannosaurids. The Kirtland Formation is interpreted as river floodplains appearing after a retreat of the Western Interior Seaway. Conifers were the dominant plants, and chasmosaurine horned dinosaurs were apparently more common than hadrosaurids.
Paleobiology
Feeding
As a hadrosaurid, Parasaurolophus was a large bipedal/quadrupedal herbivore, eating plants with a sophisticated skull that permitted a grinding motion analogous to chewing. Its teeth were continually replacing and packed into dental batteries that contained hundreds of teeth, only a relative handful of which were in use at any time. It used its beak to crop plant material, which was held in the jaws by a cheek-like organ. Feeding would have been from the ground up to around 4 meters (13 ft) above. As noted by Bob Bakker, lambeosaurines have narrower beaks than hadrosaurines, implying that Parasaurolophus and its relatives could feed more selectively than their broad-beaked, crestless counterparts.
Cranial crest
Many hypotheses have been advanced as to what functions the cranial crest of Parasaurolophus performed, but most have been discredited. It is now believed that it may have had several functions: visual display for identifying species and sex, sound amplification for communication, and thermoregulation. It is not clear which was most significant at what times in the evolution of the crest and its internal nasal passages.
Differences among species
As for other lambeosaurines, it is believed that the cranial crest of Parasaurolophus changed with age and was a sexually dimorphic characteristic in adults. James Hopson, one of the first researchers to describe lambeosaurine crests in terms of such distinctions, suggested that P. cyrtocristatus, with its small crest, was the female form of P. tubicen. Thomas Williamson suggested it was the juvenile form. Neither hypothesis became widely accepted. As only six skulls are known, additional material will help clear up these potential relationships. Williamson noted that in any case, juvenile Parasaurolophus probably had small, rounded crests like P. cyrtocristatus, that probably grew faster as individuals approached sexual maturity.
Rejected hypotheses about function
Many early suggestions focused on adaptations for a aquatic lifestyle, following the hypothesis that hadrosaurids were amphibious, a common line of thought until the 1960s. Thus, Alfred Sherwood Romer proposed it served as a snorkel, Martin Wilfarth that it was an attachment for a mobile proboscis used as a breathing tube or for food gathering, Charles M. Sternberg that it served as an airtrap to keep water out of the lungs, and Ned Colbert that it served as an air reservoir for prolonged stays underwater.
Other proposals were more physical. As mentioned above, William Parks suggested that it was joined to the vertebrae with ligaments or muscles, and helped with moving and supporting the head. Othenio Abel proposed it was used as a weapon in combat among members of the same species, and Angela Milner suggested that it could be used as a foliage deflector, like the helmet crest of the cassowary. Still other proposals made housing specialized organs the major function. Halszka Osm??lska suggested that it housed salt glands, and John Ostrom suggested that it housed expanded areas for olfactory tissue and much improved sense of smell of the lambeosaurines, which had no obvious defensive capabilities. One unusual suggestion, made by creationist Duane Gish, is that the crest housed chemical glands that allowed it to throw jets of chemical "fire" at enemies, similar to the modern-day bombardier beetle.
Most of these hypotheses have been discredited. For example, there is no hole at the end of the crest for a snorkeling function. There are no muscle scars for a proboscis and it is dubious that an animal with a beak would need one. As a proposed airlock, it would not have kept out water. The proposed air reservoir would have been insufficient for an animal the size of Parasaurolophus. Other hadrosaurids had large heads without needing large hollow crests to serve as attachment points for supporting ligaments. Also, none of the proposals explain why the crest has such a shape, why other lambeosaurines have crests that look much different but perform a similar function, how crestless or solid-crested hadrosaurids got along without such capabilities, or why some hadrosaurids had solid crests. These considerations particularly impact hypotheses based on increasing the capabilities of systems already present in the animal, such as the salt gland and olfaction hypotheses, and indicate that these were not primary functions of the crest. Additionally, work on the nasal cavity of lambeosaurines shows that olfactory nerves and corresponding sensory tissue were largely outside the portion of the nasal passages in the crest, so the expansion of the crest had little to do with the sense of smell.
Social functions
Instead, social and physiological functions have become more supported as function(s) of the crest, focusing on visual and auditory identification and communication. As a large object, the crest has clear value as a visual signal, and sets this animal apart from its contemporaries. The large size of hadrosaurid eye sockets and the presence of sclerotic rings in the eyes imply acute vision and diurnal habits, evidence that sight was important to these animals. If, as is commonly illustrated, a skin frill extended from the crest to the neck or back, the proposed visual display would have been even showier. As is suggested by other lambeosaurine skulls, the crest of Parasaurolophus likely permitted both species identification (such as separating it from Corythosaurus or Lambeosaurus) and sexual identification by shape and size.
Sounding function
However, the external appearance of the crest does not correspond to the complex internal anatomy of the nasal passages, which suggests another function accounted for usage of the internal space. Carl Wiman was the first to propose, in 1931, that the passages served an auditory signaling function, like a crumhorn; Hopson and David B. Weishampel revisited this idea in the 1970s and 1980s. Hopson found that there is anatomical evidence that hadrosaurids had strong hearing; there is at least one example, in the related Corythosaurus, of a slender stapes (reptilian ear bone) in place, which combined with a large space for an eardrum implies a sensitive middle ear, and the hadrosaurid lagena is elongate like a crocodilian's, indicating that the auditory portion of the inner ear was well-developed. Weishampel suggested that P. walkeri was able to produce frequencies of 48 to 240 Hz, and P. cyrtocristatus (interpreted as a juvenile crest form) 75 to 375 Hz. Based on similarity of hadrosaurid inner ears to those of crocodiles, he also proposed that adult hadrosaurids were sensitive to high frequencies, such as their offspring might produce. According to Weishampel, this is consistent with parents and offspring communicating.
Computer modeling of a well-preserved specimen of P. tubicen, with more complex air passages than those of P. walkeri, has allowed the reconstruction of the possible sound its crest produced. The main path resonates at around 30 Hz, but the complicated sinus anatomy causes peaks and valleys in the sound.
Cooling function
The large surface area and vascularization of the crest also suggests a thermoregulatory function. P.E. Wheeler first suggested this use in 1978 as a way keep the brain cool. Teresa Marya??ska and Osm??lska also proposed thermoregulation at about the same time, and Sullivan and Willamson took further interest. David Evans' 2006 discussion of lambeosaurine crest functions was favorable to the idea, at least as an initial factor for the evolution of crest expansion.
http://en.wikipedia.org/wiki/Parasaurolophus
| The text in this page is based on the copyrighted Wikipedia article shown in above URL. It is used under the GNU Free Documentation License. You may redistribute it, verbatim or modified, providing that you comply with the terms of the GFDL. |
|